
Indexed In
- RefSeek
- Directory of Research Journal Indexing (DRJI)
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Scholarsteer
- Publons
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
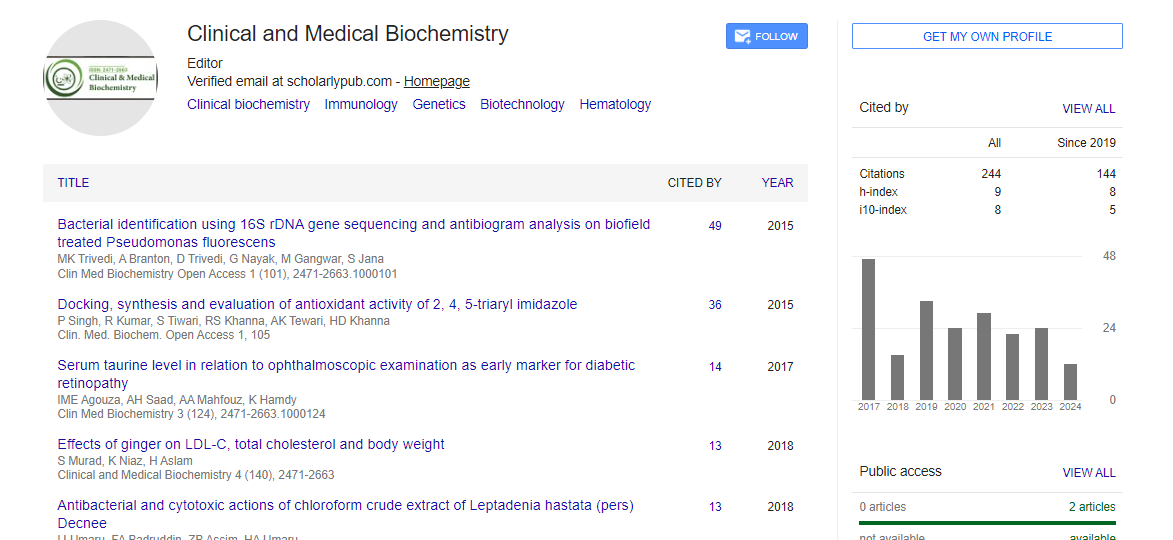
Commentary - (2023) Volume 9, Issue 2
Oxidative Stress and The Role of Thyroid Hormones in Antioxidant Regulation
Yao Duzheng*Received: 03-Mar-2023, Manuscript No. CMBO-23-20916; Editor assigned: 06-Mar-2023, Pre QC No. CMBO-23-20916 (PQ); Reviewed: 20-Mar-2023, QC No. CMBO-23-20916; Revised: 27-Mar-2023, Manuscript No. CMBO-23-20916 (R); Published: 03-Apr-2023, DOI: 10.35841/2471-2663.23.9.159
Description
Oxidative stress (OS) is characterized as an imbalance between prooxidant substance synthesis and antioxidant defenses. The reactive oxygen species (ROS) and Reactive Nitrogen Species (RNS) are the most significant prooxidants. Superoxide anion, hydroxyl radical, hydrogen peroxide, and hypochlorous acid are all members of the ROS family. During the oxidative metabolism of energy substrates, the first three compounds are primarily produced in vivo by the mitochondrial respiratory chain. RNS are peroxynitrite, which is formed when Nitric Oxide (NO) reacts with superoxide, and nitrosoperoxycarbonate, which is formed when peroxynitrite reacts with carbon dioxide. ROS and RNS are thought to be key pathogenetic variables in a variety of illnesses. Free radicals, such as superoxide anion and hydroxyl radical, have a specific pathogenetic role among them. They are molecules with high chemical reactivity due to a single unpaired electron in the exterior orbital. ROS formation may play functional functions in specific cell types, including leukocytes, endothelium and mesangial cells, fibroblasts, thyrocytes, oocytes, Leydig cells, and adipocytes. Dual Oxidases (DUOX), which generate hydrogen peroxide, are required for Thyroid Peroxidase (TPO) catalyzed hormone production.
Thyroid has two oxidases from this family (DUOX1 and DUOX2). They collaborate with DUOXA1 and DUOXA2, maturation factors that allow DUOX enzymes to translocate to the follicular cell membrane and exhibit enzymatic activity. Furthermore, NADPH Oxidase 4 (NOX4) is a novel intracellular ROS-producing mechanism recently discovered in the human thyroid gland. Increased ROS production by the respiratory chain as a result of increased energetic demand or substrate availability, as seen in obesity, or mitochondrial dysfunction or impairment, can cause cell damage and contribute to the pathophysiology of various diseases, including inflammatory (e.g., rheumatoid arthritis) and cardiovascular (e.g., myocardial infarction). A pathogenic function for ROS has also been proposed in diabetes mellitus, where oxidation occurs concurrently with glycation in vivo and antioxidant capacity is reduced, resulting in greater sensitivity to oxidative stress. The endoplasmic reticulum, mitochondria, plasma membrane, peroxisomes, and cytoplasm have all been found to have different defensive mechanisms that guard against free radical damage. Free radicals are prevented or promptly inactivated by enzymes such as Superoxide Dismutase (SOD), Catalase (CAT), and Glutathione Peroxidase (GPx), as well as transition-metal binding proteins such as transferrin, ferritin, and ceruloplasmin.
SOD speeds up the dismutation of superoxide anion in hydrogen peroxide and molecular oxygen, which normally takes 104 times longer. By converting hydrogen peroxide into water and molecular oxygen, CAT detoxifies it. When hydrogen peroxide levels are high, GPx also participates in detoxification. Furthermore, GPx detoxifies lipid peroxides by converting them to the appropriate alcohols. "Scavenger" molecules, which include both water-soluble and liposoluble molecules such as albumin, bilirubin, ascorbic acid, urates, and thiols, interrupt the lipid-peroxidation process by interacting with and neutralizing the intermediate radicals. Scavengers have a rapid diffusion rate in biological membranes, allowing them to intercept radicals and convert them into more stable molecules, effectively halting the radical chain. Scavengers can sometimes be regenerated. A third defence strategy employs procedures that remove the molecules destroyed by the oxidative attack, allowing normal structures to be reconstituted. In response to metabolic overload caused by an abundance of macronutrients, ROS and RNS can be produced at the cellular level. Moreover, mitochondrial dysfunction and endothelial reticulum stress lead to metabolic abnormalities in adipose tissue in obese patients. Reduced glutathione (GSH) is an essential cofactor for both antioxidant enzymes and deiodinases, which convert Thyroxine (T4) to Triiodothyronine (T3). Furthermore, plasma levels of tiny antioxidant molecules including Vitamin E and CoQ10, as well as thyroid hormones, are linked. OS has been associated to both hyperthyroidism and hypothyroidism, with autoimmune thyroiditis or a functional picture of low-T3 syndrome found in acute and chronic Nonthyroidal Disease Syndrome (NTIS). In the context of reproductive endocrinology, the role of thyroid in the control of antioxidant systems has recently been addressed.
Thyroid function is well established to influence ovarian activity. ROS play physiological roles in the ovary, and hypothyroidism (low T3) can wreak havoc on the ovary by interfering with antioxidant systems. OS has been linked to both hyperthyroidism and hypothyroidism. However, the mechanisms by which OS is produced differ in these two clinical scenarios: increased ROS production in hyperthyroidism and decreased antioxidant availability in hypothyroidism. OS causes some hyperthyroidism problems in target tissues. Thyroid hormones, unlike CAT, can act as oxidants and cause DNA damage, most likely via the phenolic group, which is comparable to that of steroidal oestrogens. Numerous other processes, as previously discussed, may be involved, including increased Nitric Oxide Synthase (NOS) gene expression with NO overproduction and activation of hepatic NF-kB with the resulting increase in cytokine levels, which stimulates ROS production. Some thyroid hormone-regulated processes, on the other hand, use autoloop feedback to fine-tune the oxidative status. We highlight the roles of Uncoupling Protein -2 (UCP) and Uncoupling Protein-3. These compounds have antioxidant activity in plants and animals, according to data. Nevertheless, only T3 appears to control UCP, while T4 has little effect. Estrogens, on the other hand, promote ROS generation by suppressing UCP. Increased mitochondrial protein turnover and mitoptosis also contribute to the regulation of oxidative state by eliminating OS-damaged mitochondria. These processes are controlled by peroxisome proliferator-activated receptor gamma coactivator-1, which is activated by T3. Thyroid hormones affect the lipid content of rat tissues and, as a result, susceptibility to OS. However, the response is tissue-specific, and disparities in T3 and T4 effects have been described. T3-induced hyperthyroidism was linked to altered lipid-peroxidation indexes in rat liver, including higher levels of Thiobarbituric Reactive Substances (TBARS) and lipid hydroperoxides, which are consequences of lipid peroxidation.
Citation: Duzheng Y (2023) Oxidative Stress and the Role of Thyroid Hormones in Antioxidant Regulation. Clin Med Bio Chem. 9:159.
Copyright: © 2023 Duzheng Y. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.