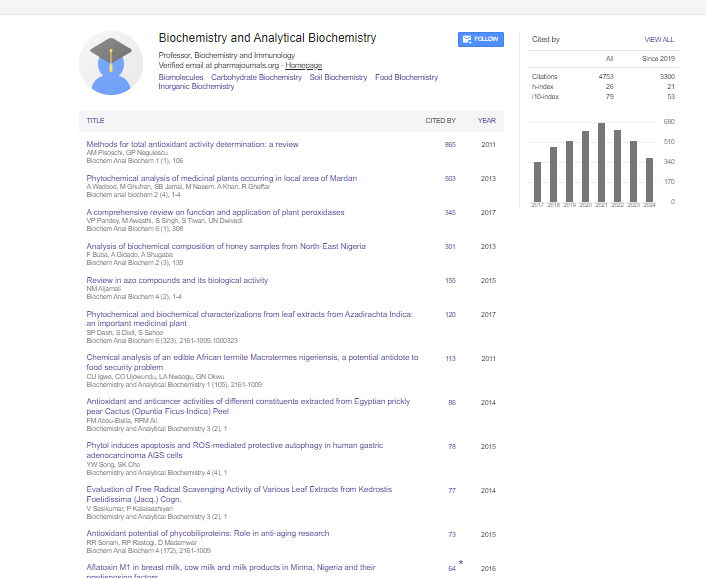


Indexed In
- Open J Gate
- Genamics JournalSeek
- ResearchBible
- RefSeek
- Directory of Research Journal Indexing (DRJI)
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Scholarsteer
- Publons
- MIAR
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Perspective - (2022) Volume 11, Issue 4
Methylation of Lysine Signals on Non-Histone Proteins.
Received: 31-Mar-2022, Manuscript No. BABCR-22-16610; Editor assigned: 04-Apr-2022, Pre QC No. BABCR-22-16610(PQ); Reviewed: 19-Apr-2022, QC No. BABCR-22-16610; Revised: 26-Apr-2022, Manuscript No. BABCR-22-16610(R); Published: 03-May-2022, DOI: 10.35248/2161-1009-22.11.429
About the Study
The study of protein structure and function at a vast scale, known as proteomics, has tremendously aided our understanding of cellular biology and disease. Through minor chemical changes, it has become clear that the proteome is geographically, chronologically, and chemically dynamic, allowing the same protein to execute significantly varied activities and fulfill completely unrelated tasks in a cell. This is known as epiproteomics, which effects epigenetic alterations to DNA that are not encoded in the DNA sequence and mostly encompasses Post-Translational Modifications (PTMs) such as phosphorylation, ubiquitination, acetylation, and methylation, which number in the hundreds. These differ based on cell type, signalling, stress, microenvironment, and other factors, resulting in changes in protein characteristics that, either directly or indirectly, affect cell function.
Proteinlysinemethyl transferase catalase lysine methylation, a key post translational alteration that regulates several signalling pathways. It affects chromatin shape and transcription both directly and indirectly. Increasing data reveals that PKMT activity dysregulation plays a key role in the development of many diseases. While the majority of these studies focus on methylation events in the context of histones, it has recently become clear that methylation of non-histone proteins is also important in cell functions. Methylation of lysine in non-histone proteins is one of the PTMs that are attracting increased interest for a variety of reasons. Protein lysine methyltransferases catalyze mono, di and tri methylation of lysine's epsilon amine (PKMTs). Initially, lysine methylation was researched extensively on a variety of histone sites, with key functions in chromatin packing and gene transcription control (both activation and repression). It has now been discovered to serve a variety of critical roles in non-histone proteins which are mostly transcription factors and chaperones via influencing function, Protein stability or half-life, as well as interaction strength with target proteins or DNA. Furthermore, because lysine can be altered in a variety of ways, methylation might compete with other PTMs, giving another layer of control.
The tumour suppressor p53 is a notable example of this, as methylation of different lysine residues or varying degrees of methylation whether mono or di leads to changes in characteristics ranging from suppressing gene transcription to increasing affinity for 53BP1. Recently, two fascinating instances have been discovered. The first is the discovery of Heat Shock Protein 70 (HSP70) acting as a transcription factor after lysine methylation, translocation to the nucleus, and increased Aurora kinase B (AURKB) kinase activity, and the second is the isolation of a group of PKMTs that do not act on histones but appear to act primarily on transcription factors and chaperones. SUV39H1 and EZH2 was shown to be upregulated in a variety of human cancers, and a gain-of-function mutation in EZH2 has been shown to increase catalytic activity and contribute to carcinogenesis while truncating mutations in METTL23 have been linked to Intellectual Disability (ID).
Despite the fact that lysine methylation is important in cellular biology and human disease, little is known about target proteins, residue position, degree, enzyme, and function. The absence of dependable instruments and methodologies for studying these alterations is one of the key reasons for this. Top-down proteomic approaches allow for protein sequencing and PTM identification; however the mass spectrometry equipment employed must have a high resolution. The addition of a phosphate group to a protein raises the mass by 80 Da (and gives an overall negative charge) or an acetyl group adds 42 Da (and neutralises the positive charge on the lysine residue), but the addition of a methyl group adds just 14Da (and does not change the charge). As a result, the change in mass on charge (m/z) after methylation is extremely tiny. Another stumbling block is the scarcity and poor quality of commercial lysine methylation antibodies.
Conclusion
The excellent thing is that these are merely temporary setbacks. Both in terms of enhanced resolution, especially when using Electron Transfer Dissociation (ETD), and in the development of enrichment procedures that improve the stoichiometric presence of changed species, technological improvements in mass spectrometry are highly rapid. Antibodies, on the other hand, are continually being made and tested. As research interest deepens, more vigorous and dependable anti-methyl lysine antibodies would become accessible, just as anti-phospho and antiacetyl antibodies developed. As with phosphorylation, lysine methylation could eventually be included in conventional diagnostic tests and targeted therapy.
Citation: Ren C (2022) Methylation of Lysine Signals on Non-Histone Proteins. Biochem Anal Biochem. 11:429.
Copyright: © 2022 Ren C. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.