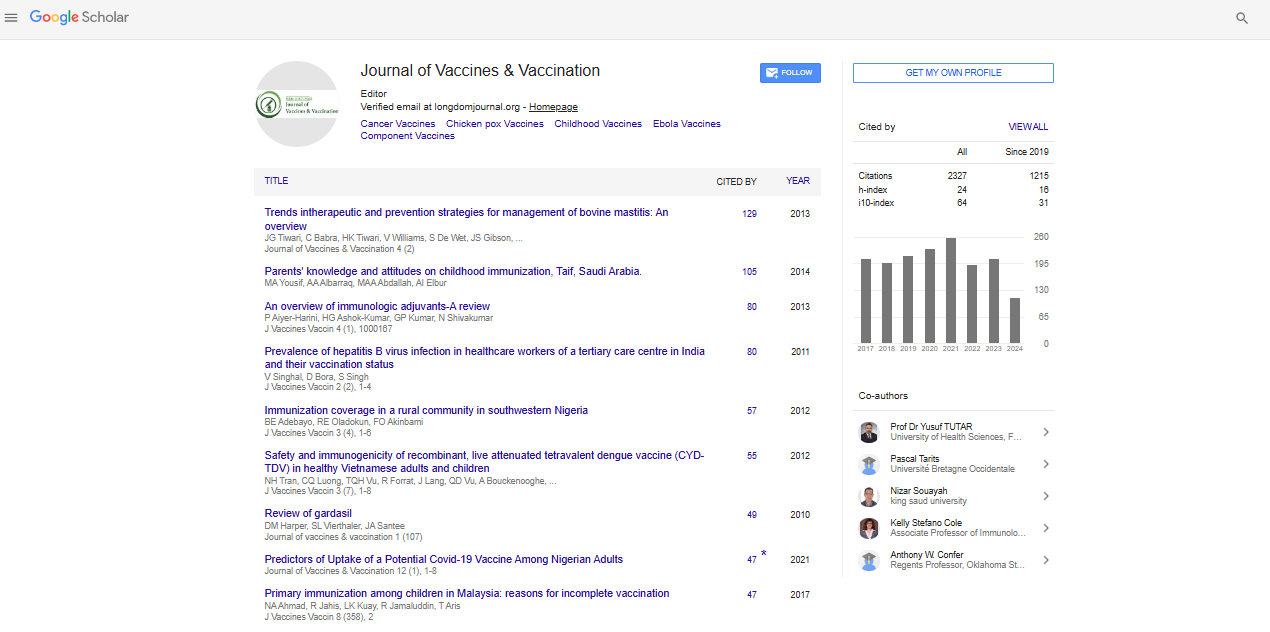

Indexed In
- Academic Journals Database
- Open J Gate
- Genamics JournalSeek
- JournalTOCs
- China National Knowledge Infrastructure (CNKI)
- Scimago
- Ulrich's Periodicals Directory
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- MIAR
- University Grants Commission
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Research Article - (2020) Volume 0, Issue 0
Assessment of the Immunogenicity of a Novel Live Recombinant Rift Valley Fever arMP- 12ΔNSm21/384 Vaccine Candidate Following Intranasal Vaccination of Goats, Sheep and Calves in Tanzania
Nyundo S.1, Adamson E.K.1, Rowland J.2, Palermo P.M.2, Salekwa L.1,4, Matiko M.K.1, Bettinger G.E.2, Wambura P.1, Morrill J.C.3, Sullivan T.R.5 and Watts D.M.2*2Department of Biological Sciences, El Paso, Texas 79968, USA
3Orion Research & Management Services Inc., 205 FM 107, Gatesville, Texas 76528, USA
4Department of Medical Sciences and Technology, Mbeya University of Science and Technology, P.O.Box 131, Mbeya, Tanzania
5Zeteo Biomedical LLC, PO Box 342216, Austin, TX 78734, USA
Received: 22-Sep-2020 Published: 14-Oct-2020, DOI: 10.35248/2157-7560.20.S5:002
Abstract
Background: Rift Valley fever virus (RVFV) is an arbovirus that causes morbidity and mortality in livestock and humans throughout Africa and in the Arabian Peninsula. Vaccines are effective for the prevention of Rift Valley fever (RVF) disease, but new and improved vaccines are needed to improve the safety of available vaccines. Also, non-invasive needle free vaccine delivery routes should be evaluated as an alternative for invasive routes of vaccination.
Objective: The aim of this proof of concept study was to evaluate the safety and immunogenicity of a novel live attenuated recombinant RVFV arMP-12ΔNSm21/384 vaccine candidate following intranasal vaccination of goats, calves, and sheep in Tanzania.
Methods: Healthy, 6-9 months old breeds of African sheep (Ovis aeris), goats (Capra aegagrus) and zebu calves (Bos taurus indicus) were used in this study. The animals were purchased from local livestock keepers in the Mvomero district of Morogoro region, Tanzania. Animals were seronegative to both RVFV and antibody at the time of use in the vaccine trials. Animals in the test group included 10 goats, 7 sheep and 10 calves that were vaccinated in the left nares with 50 μl each and 2 sheep were vaccinated with 100 μl each (50 μl each in the left and right nares) of a dose that contained 4 × 105 PFU/50 ul of arMP-12ΔNSm21/384 vaccine, while the control group, including 2 goats, 3 sheep and 2 calves that were injected in the left nares with 50 μl of phosphate buffered saline to serve as placebo controls. Rectal temperature was measured and blood samples were collected on day 14 and 0 before vaccination, and on days 3, 5, 7, 14, 21, 28 and 35 post vaccinations (PV). Serum samples collected on days 14 and 0 before vaccination were tested for RVFV neutralizing antibody by a plaque reduction neutralization test, and on days 3 and 5 PV, serum samples were tested for virus as possible evidence of a viremia in cell culture and weekly collected samples thereafter were tested for RVFV neutralizing antibody.
Results: All animals were negative for RVFV neutralizing antibody at 14 and 0 days before vaccination and none of the animals had detectable viremia on days 3 and 5 PV, and none had clinical manifestations throughout the study. Among the 7 sheep, 10 goats, and 10 calves that received 50 μl each of the vaccine dose, 70% had the first detectable antibody on either day 5, 7 or 14 PV with titers ranging from 1:10 to 1:40. The 2 sheep that received the 100 μl each of the virus dose had the first detectable antibody on day 5 PV with a titer of 1:160. Subsequently, animals vaccinated with the 50 μl dose had antibody titers ranged from 1:10 to 640 on days 21, 28 and 35 PV, while those vaccinated with 100 μl maintained an antibody titer of 1:160 throughout the study. Moreover, there was no difference in the antibody titers between animal species p=0.34, although mean antibody titers of goats were highest.
Conclusion: As a proof of concept studies, the findings demonstrated that intranasal vaccination is a promising route for vaccinating domestic ruminants with the RVFV arMP-12ΔNSm21/384 vaccine candidate. However, these preliminary results suggested that a larger dose of 4 × 105 PFU/100 ul of arMP-12ΔNSm21/384 vaccine needed to be administered to each animal to consistently elicit a robust immune response. Also, further studies are warranted using a larger number of domestic ruminants to confirm the immune response elicited by the larger dose of the vaccine administered via the intranasal route.
Keywords
Rift valley fever (RVF); RVF virus; RVF arMP-12ΔNSm21/384 vaccine; RVF antibody; Intranasal vaccination; Goat; Sheep; Calves; Tanzania
Introduction
Rift valley fever (RVF) is an acute viral zoonotic disease caused by RVF virus (RVFV) that affects both ruminants and humans. The disease was first characterized by Daubney and his co-worker in 1934 during an outbreak of RVF in Kenya among exotic wool sheep that had been imported into East Africa [1]. Outbreaks of RVF were confined to the Rift valley of East Africa until 1977 when a major outbreak occurred in Egypt [2]. The virus has since been documented to be enzootic throughout much of sub-Saharan Africa [3-5] and in 2000, the virus was first reported outside Africa as the cause of an outbreak in Saudi Arabia and Yemen and therefore, considered to have the potential for global spread [6,7]. More RVF outbreaks were reported during 2003 in Egypt, 2006- 2007 in Kenya, Somalia and Tanzania, 2007 in Sudan, 2018-19 in Madagascar, 2010 in South Africa, 2012 Mauritania, and 2016 in Niger [8-13].
Susceptibility to RVFV infection depends on the age and animal species. In young animals, RVFV causes mortality of up to 100% and abortions in pregnant ewes of approximately 80%-100% [14]. Clinical signs in adult sheep, goats and cattle are not consistent but may include a rise in body temperature, nasal discharge, unsteady gait, excessive salivation, loss of appetite, and bloody diarrhea [13,15]. High abortion rates alone among animals with a febrile illness can be a strong indicator of a RVFV outbreak, especially following periods of heavy rainfall in east Africa when the mosquito vectors appears in large numbers [4,16].
In humans, RVF disease presents as a mild febrile illness that may progress to severe illness characterized by fever, dizziness, weight loss and myalgia and in some patients, disease may advance to severe hemorrhagic fever, encephalitis, and ocular disease [17-21]. Mortality rates usually range from 1%-4%, although some people recover within 4-7 days and some progressing to severe and fatal disease [13]. The RVFV is mainly transmitted by mosquito vectors of Aedes species. However, the virus can be transmitted to humans through direct contact with RVFV contaminated fluid and tissue material [21-24]. Exposure to infectious aerosols has been confirmed as a source of RVFV infection [25]. Also, RVFV was observed to be stable in tap water for several days and mice were susceptible to infection through drinking RVFV contaminated water [26].
RVFV virus belongs to the genus Phlebovirus, family Phenuiviridae, a group of enveloped RNA viruses [27]. The enveloped virion is about 100 nm in diameter with glycoproteins protruding from the surface [28]. It is spherical in shape and composed of single stranded RNA that is surrounded by nucleocapsid proteins (NP). The RNA genome is divided into three segments each attached with RNA dependent RNA polymerase [29-31]. The segments include the small (S), medium (M) and large (L) RNA of 1.6 kb, 3.8 kb and 6.4 kb, respectively. The S segment is 1690 nucleotides long [31,32] and encodes a nucleocapsid protein in the negative sense and a non-structural protein (NSs) in a positive sense [33,34]. The NP is highly immunogenic and many antibody-based RVFV detection assays are based on this protein.
Among attempts to control RVF in animals and humans, vaccines have proven to be most effective. For veterinary use, there are two live attenuated vaccines, the Smithburn neurotropic and Clone-13, that have been used in previous RVF outbreaks [35]. However, the Smith-burn vaccine causes abortions and malformations in pregnant ewes similar to the wild type RVFV and the Clone 13 is temperature sensitive with the potential to cause teratogenic effect among pregnant sheep based on experimental studies [36,37]. In addition, both vaccines elicit an immune response that does not distinguish infected from vaccinated animals (DIVA). DIVA is important in avoiding trade restriction and exportation of infected animal in Africa [38]. Currently, there are no approved vaccines for human use [35]. The MP-12 strain used in deriving MP-12ΔNSm21/384 vaccine candidate has been tested in humans, sheep, cattle and non-human primates through subcutaneous or intramuscular route and found to be safe and immunogenic [26,39,40]. However the vaccine is not DIVA compatible. As an approach to develop a DIVA compatible vaccine, the MP-12 vaccine virus was used to develop a recombinant vaccine using reverse genetics technology to delete nucleotides 21-384 of the NSm gene from the M segment of the MP-12 virus strain to produce the RVFV MP-12ΔNSm21/384 candidate vaccine to serve as a potential DIVA vaccine [41-45]. Subsequent studies showed the RVFV arMP- 12ΔNSm21/384 vaccine candidate to be safe, and immunogenic in sheep and calves [45,57,63] and efficacious in sheep against a challenge dose of virulent RVFV ZH-508 [63]. Studies have shown that many viruses attach to the mucus membrane of the oral or nasal cavity, and elicit a protective immune response at local and distal sites [46,47,48]. MP-12 has been shown to immunize monkeys by aerosol and also provide protection against virulent RVFV against aerosol infection [53], and supports the evaluation intranasal immunization of livestock with MP-12ΔNSm21/384.
Needle injection is the major method for vaccination and drug delivery to livestock in order to induce immune response using safe and highly purified vaccines. Although needle and syringe devices are inexpensive and easily adaptable to different settings, needle free technology offers advantages as compared to needle free delivery of vaccine [49,50]. The needle free vaccine delivery technology enhances safety, and delivers vaccine/drug without piercing the skin that results in fewer injection site lesions [49,50]. Therefore, there is a benefit to change from needle based to needle free delivery. Currently an increasing number of influenza vaccines studies have confirmed the efficacy of the needle free delivery of vaccines using different routes of administration such as intranasal and intradermal routes [51]. This study was conducted to evaluate the safety and immunogenicity of RVFV MP-12ΔNSm21/384 vaccine candidates in Ovis aries sheep, Bos taurus indicus zebu calves and Capra aegrus goats using an experimental intranasal vaccine aerosol dispenser.
Materials and Methods
Study area
The present study was conducted in Morogoro, Tanzania (6.8278°S, 37.6591°E) at Sokoine University of Agriculture (SUA) in a vector controlled Animal Biosafety Level 2 (ABSL-2) facility and a Biosafety Level 2 (BSL-2) virology laboratory. The animal facility is designed to prevent the entry of arthropods and to provide sanitation measures as well as being equipped with incinerator for disposing animal animal waste. The laboratory is equipped with basic virology and cell culture instruments together with a distiller and an autoclave for autoclaving equipment before use and autoclaving waste materials from the laboratory before exposing the waste to the environment.
Experimental animals
Healthy, 6-9 months old sheep (Ovis aeris), goats (Capra aegagrus) and zebu calves (Bos taurus indicus) were used in this study. A total of 36 animals for intranasal injection (12 goats, 12 sheep and 12 zebu calves) were purchased from local livestock keepers at Mvomero district of Morogoro region, Tanzania. The animals were transported to a temporary animal holding facility located near the permanent animal housing in the SUA Animal Biosafety Laboratory (ABSL)-2 facilities. They were ear tagged with individual identification numbers and treated with Steladone 300® ECAcaricide, and given 2.5% Albendazole orally in order to remove ectoparasites and endoparasites respectively. Animals were then held for two weeks to acclimatize. Throughout the experiment, the animals were fed “ad libitum” with fresh grasses, water, and mineral blocks. They were monitored daily for elevated body temperature to detect any illnesses. The animals were then moved into the ABSL-2 facility. All animals were managed and treated in accordance with a protocol approved by the Institutional Animal Care and Use Committee approved by University of Texas at El Paso (UTEP), Texas and SUA Institutional Animal Care and Use Committee (IACUC) (ref # 559105-08 and SUA/CMVBS/R.1).
Vero E6 cells and vaccine viruses
The Vero E6 cells used in this study were provided by the University of Texas at El Paso (UTEP), Texas. The RVFV MP-12 virus was originally obtained by UTEP from the World Reference Centre for Emerging Viruses and Arboviruses, Department of Microbiology and Immunology, University of Texas Medical Branch, Galveston Texas. At UTEP, the identity of the RVF MP-12 vaccine virus was confirmed by plaque reduction neutralization test (PRNT) using RVF MP-12 specific monoclonal antibody (Mab). The Mab neutralized the infectivity titer of the RVF MP-12 virus from 106 plaque forming units (PFU)/ml to 102 PFU/ml, but did not neutralize the infectivity titer of Sindbis and/or West Nile viruses.
The RVFV MP-12 obtained from UTEP was used at SUA to prepare a stock virus in Vero E6 cells with an infectivity titer of 1.4 × 107 PFU/ml and stored in 0.5 ml aliquots at -80°C for use to perform the Plaque Reduction Neutralization Test (PRNT) to detect RVFV neutralizing antibody in sera samples obtained vaccinated animals.
A stock virus of RVF arMP-12ΔNSm21/384 with an infectivity titer of 8 × 106 PFU/ml was prepared by passaging in Vero cells. The vaccine for intranasal delivery was prefilled in a volume of 50 μl in each unit dose blister (packed in a delivery cartridge tip) by Mystic Pharmaceuticals, Inc., Austin, Texas and stored at -80°C.
Experimental design
The calves, sheep and goats were divided into two groups, including twenty seven animals, 10 goats, 7 sheep and 10 zebu calves that were vaccinated in the left nares with 50 μl each of 4 × 105 plaque forming units (PFU) with the RVFV arMP-12ΔNSm21/384 vaccine. A second group of 7 animals, 2 goats, 3 sheep and 2 zebu calves received 50 μl of phosphate buffered saline (PBS) to serve as negative controls. Also, two sheep were inoculated with a double volume (100 μl) via the intranasal route (8 × 105 PFU) of the arMP- 12ΔNSm21/384 vaccine.
All animals were vaccinated using Mystic’s large animal nasal delivery device. As a mechanical dispensing device, the components consisted of a threshold force actuated trigger and plunger designed to generate a consistent actuation force and a blister packed in an individual nasal cartridge tip designed to fit in the nares of each animal species. Each blister contained an internal micro-nozzle designed to aerosolize the vaccine to droplets ranging from 50 to 100 microns to facilitate deposition to the nasal mucosal surfaces. The blisters were pre-loaded with the desired volume and concentration of the vaccine and then sealed leaving a small amount of compressible head space inside each blister. The vaccine was then dispensed by pressing the trigger to push the plunger to compress and crush the blister expelling the vaccine through the internal nozzle which emerges from inside the blister and extends out of an orifice in the cartridge tip dispensing the vaccine as a fine spray into the animal’s nasal cavity to penetrate the surrounding mucosal tissue.
Specimen collection and preparation
Blood samples of 4 ml were collected from the jugular vein of all animals in phase I and phase II using a 6 ml vacutainer 14 days before vaccination, and on day 0 immediately before vaccination and were tested for RVFV by cell culture assay and RVFV antibody by PRNT. Samples obtained on days 3, 4, and 5 were tested for RVFV by cell culture assay, and thereafter, samples obtained on days 7, 14, 21, 28 and 35 post vaccination (PV) were tested to determine the neutralizing antibody response by the PRNT. Two to 3 ml of serum was obtained from each blood sample after leaving the samples overnight at 4°C followed by centrifugation at 1200 × G for 10 minutes. Aliquots of 0.5 to 1.0 ml of each serum sample were transferred to sterile pre-labeled vials and stored at -80°C freezer until tested for RVFV and/or RVFV neutralizing antibody.
Virus isolation
Test animal serum samples were diluted in 1:2 in Eagle's Minimum Essential Medium (EMEM) supplemented with 4% Fetal Bovine Serum (FBS). Confluent monolayer of Vero E6 cells were propagated in 24 well plates and cultures in each well were inoculated in duplicate using 50 μl of each serum sample per culture. The culture and inoculum were incubated for one hour at 37°C and agitated after every 15 minutes for virus absorption. After absorption, 0.5 ml of EMEM supplemented with 4% FBS was added to each culture and incubated at 37°C with 5% CO2. Cultures were observed once daily for 10 days under microscope for cytopathic effect (CPE). After 10 days, all CPE negative cultures were frozen and thawed to make a blind passage using the same procedure and observed for 10 days for CPE. Any cultures that developed CPE were harvested and stored in aliquots of 1.0 ml for further study using RT-PCR to determine if the CPE was caused by RVFV. If there was presumptive evidence of wild type RVFV, all aliquots and any remaining cultures would be destroyed by autoclaving. The specific animals would be isolated and quarantined in a holding facility separate from the ABSL 2 facility and not used in this study.
Immunological method
Plaque reduction neutralization test-: Each test serum was diluted 1:5 followed by 4-fold dilutions of 1:5, 1:20, 1:80, 1:320, 1:1280 and 1:5120 in Hanks Balanced Salt Solution (HBSS) supplemented with 1% HEPES, penicillin and streptomycin and heat inactivated FBS in a 96-well plate. An equal volume of 75 μl of each diluted test sera was mixed with an equal volume of 75 μl of RVFV MP- 12 with approximately 60-80 PFUs such that the final serum dilution were 1:10, 1:40, 1:160, 1:640, 1:2560 and 1:10240, and virus dose ranges from 30 to 40 PFU. The controls consisted of a mixture of equal volume of 60-80 PFU with a 1:10 dilution of RVFV antibody positive and a negative goat serum. The virus dose serum dilution mixtures were incubated at 37°C in the absence of CO2 for one hour. Next, Vero E6 cells were seeded in 24-well tissue culture plates and incubated for 2-3 days at 37°C and 5% CO2 to provide about 90% confluence monolayers. The growth media was then discarded from the Vero cell monolayers and 50 μl of each virus dose-serum dilution mixture was inoculated onto each of 2 cell monolayers per sample. The mixture of the virus dose and the antibody positive control serum mixture were inoculated onto each of 20 cultures and the virus dose-antibody negative control serum mixture was inoculated onto 4 cultures Plates were incubated for one hour at 37°C and 5% CO2 while agitating after every 15 minutes. One percent Seakem agarose with an equal volume of 2x Eagle’s Basal Medium with Earle’s salt (EBME), with HEPES, sodium bicarbonate, 8% FBS, and 1% penicillin, streptomycin and L-glutamine was prepared, and 0.5 ml was overlaid onto cell culture of each well. The agarose overlay was allowed to solidify and plates with cells were incubated for 3 days at 37°C with 5% CO2. Each culture of cells and inoculum was then overlaid with 0.5 ml of 1% agarose mixed with an equal volume of 2x EBME supplemented with 5% neutral red and incubated overnight at 37°C. The dilution of serum that reduced the RVF MP-12 virus dose by 80% was considered as the neutralizing antibody titer.
Post vaccination observation
Rectal body temperature for each animal was taken at the time of blood collection up to day 35 PV. In addition, general health status was assessed by veterinary personnel and recorded once a day. Animals that developed any sign of illness during the study were given a clinical examination by a veterinarian and samples were collected for analysis and diagnosis.
Statistical analysis
Data analysis were done using R statistical analysis software version 3.4.1. Analysis of mean PRNT 80 antibody titers among animal species were done using welch two sample t-test and One-way ANOVA with a significance level of p ≤ 0.05.
Results
Post vaccination observations
The average body temperatures for calves, goats and sheep vaccinated IN with arMP-12ΔNSm21/384 as well as of negative control animals vaccinated with PBS are presented in Figures 1(a)- 1(c).

Figure 1a: Mean rectal temperature of calves inoculated intranasally with 4 × 105 PFU/ml of arMP-12∆NSm21/ 384 vaccines and phosphate buffered saline (PBS).

Figure 1b: Mean rectal temperature of sheep inoculated intranasally with 4 × 105 PFU/ml of arMP-12∆NSm21/ 384 vaccines and phosphate buffered saline (PBS).

Figure 1c: Mean rectal temperature of goats inoculated intranasally with 4 × 105 PFU/ml of arMP-12∆NSm21/ 384 vaccines and phosphate buffered saline (PBS).
Normal body temperatures were observed that ranged from 37.2°C to 39.8°C in the arMP-12ΔNSm21/384 vaccinated animals and 37.5°C to 39.5°C in control animals. None of the animals developed clinical signs of illness such as fever throughout the study.
Viremia
A RVFV viremia was not detected in any of the animals before and after they were vaccinated with arMP-12ΔNSm21/384 or among the control animals that received PBS. This observation was supported by unsuccessful attempts to isolate RVFV virus from sera samples obtained from goats, sheep and calves at 14 days before vaccination and on day 0 immediately prior to vaccination and on days 0, 3, 4 and 5 PV.
Immunogenicity
A total of 70% of the calves, sheep and goats vaccinated via the IN route with 50 μl each for the arMP-12ΔNSm21/384 developed detectable neutralizing antibody. Neutralizing antibody was observed beginning on day 5 PV in 3 of 7 calves and 1 of 9 goats (#979). None of the sheep had detectable neutralizing antibody on day 5 PV but antibody was detected in all sheep on day 7 and 14 PV together with the remaining goats and calves that were negative on day 5. The antibody titers of the vaccinated animals with 50 μl ranged from 1:10 to 1:40 in calves, 1:10 to 1:40 in sheep and 1:10 to 1:640 in goats. A total of 9 vaccinated animals, 3 calves (#911, #918 and #902), 3 sheep (#938, #942 and #949) and 3 goat (#958, #965 and #969) did not develop detectable antibody through day 35, thus resembling PBS vaccinated control animals that did not have detectable neutralizing antibody throughout the study (Table 1).
| Days post vaccination | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Vaccine | Volume | Animal # | -14 | 0 | 5 | 7 | 14 | 21 | 28 | 35 |
| Calf | PBS | 0.05 | 913 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calf | PBS | 0.05 | 923 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | PBS | 0.05 | 943 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | PBS | 0.05 | 951 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | PBS | 0.05 | 954 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Goat | PBS | 0.05 | 974 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Goat | PBS | 0.05 | 980 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calf | arMP12∆NSm | 0.05 | 911 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calf | arMP12∆NSm | 0.05 | 921 | 0 | 0 | 10 | 10 | 10 | 10 | 10 | 10 |
| Calf | arMP12∆NSm | 0.05 | 928 | 0 | 0 | 0 | 10 | 40 | 40 | 40 | 40 |
| Calf | arMP12∆NSm | 0.05 | 918 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 |
| Calf | arMP12∆NSm | 0.05 | 902 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calf | arMP12∆NSm | 0.05 | 908 | 0 | 0 | 0 | 0 | 10 | 10 | 10 | 10 |
| Calf | arMP12∆NSm | 0.05 | 926 | 0 | 0 | 0 | 0 | 0 | 10 | 10 | 10 |
| Calf | arMP12∆NSm | 0.05 | 915 | 0 | 0 | 10 | 10 | 40 | 40 | 40 | 40 |
| Calf | arMP12∆NSm | 0.05 | 907 | 0 | 0 | 10 | 10 | 10 | 10 | 10 | 10 |
| Calf | arMP12∆NSm | 0.05 | 924 | 0 | 0 | 0 | 0 | 10 | 40 | 40 | 40 |
| Sheep | arMP12∆NSm | 0.1 | 930 | 0 | 0 | 160 | 160 | 160 | 160 | 160 | 160 |
| Sheep | arMP12∆NSm | 0.05 | 933 | 0 | 0 | 10 | 10 | 10 | 10 | 10 | 10 |
| Sheep | arMP12∆NSm | 0.1 | 935 | 0 | 0 | 160 | 160 | 160 | 160 | 160 | 160 |
| Sheep | arMP12∆NSm | 0.05 | 938 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | arMP12∆NSm | 0.05 | 939 | 0 | 0 | 0 | 0 | 40 | 40 | 40 | 40 |
| Sheep | arMP12∆NSm | 0.05 | 942 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | arMP12∆NSm | 0.05 | 949 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sheep | arMP12∆NSm | 0.05 | 955 | 0 | 0 | 0 | 0 | 40 | 40 | 40 | 40 |
| Sheep | arMP12∆NSm | 0.05 | 957 | 0 | 0 | 0 | 0 | 0 | 10 | 10 | 10 |
| Goat | arMP12∆NSm | 0.05 | 968 | 0 | 0 | 0 | 10 | 10 | 10 | 10 | 10 |
| Goat | arMP12∆NSm | 0.05 | 958 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Goat | arMP12∆NSm | 0.05 | 982 | 0 | 0 | 0 | 0 | 40 | 10 | 10 | 10 |
| Goat | arMP12∆NSm | 0.05 | 977 | 0 | 0 | 0 | 0 | 40 | 40 | 40 | 40 |
| Goat | arMP12∆NSm | 0.05 | 961 | 0 | 0 | 0 | 0 | 40 | 160 | 160 | 160 |
| Goat | arMP12∆NSm | 0.05 | 965 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Goat | arMP12∆NSm | 0.05 | 979 | 0 | 10 | 10 | 10 | 10 | 640 | 160 | 160 |
| Goat | arMP12∆NSm | 0.05 | 976 | 0 | 0 | 0 | 0 | 10 | 10 | 40 | 40 |
| Goat | arMP12∆NSm | 0.05 | 984 | 0 | 0 | 0 | 0 | 160 | 160 | 40 | 40 |
| Goat | arMP12∆NSm | 0.05 | 969 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Table 1: Rift valley fever virus neutralizing antibody titers in calves, sheep and goats vaccinated (4 × 105-8 × 105 PFU) of arMP-12∆NSm21/384 vaccine and phosphate buffered saline (PBS).
The two sheep that received the 100 μl of the vaccine had an antibody titer of 1:160 that was first detected on day 5 and the titer was sustained through day 35 PV. The antibody mean titers among goats, sheep and calves vaccinated with arMP-12ΔNSm21/384 did not differ significantly (p=0.34). However, goats had slightly higher antibody titers than sheep and calves.
Discussion
The overall aim of this study was to determine the safety and immunogenicity of the RVFV arMP-12ΔNSm21/384 vaccine candidate in economically and food source susceptible livestock species, using the IN route of administration. Viremia was not detected in any of the vaccinated and control animals which demonstr potential transmission by mosquito vectors [54-56]. Moreover, there was no inflammatory reaction at the exposure site in all vaccinated animals using the Mystic vaccine delivery dispenser. Only minimal skills are required to use this device, thus avoiding the use of needles that can result in needle stick injuries and accelerate virus transmissions during the vaccination of animals. None of the animals developed any signs and/or symptoms of illness, thus confirming the safety of the vaccine in terms of causing fever or other clinical manifestations among domestic ruminants [57-61]. The rectal temperatures of both controls and vaccinated animals was approximately 39°C at the time the animals were vaccinated and then stabilized to range from 37.5°C to 38.5°C throughout the study. The slightly elevated temperature was probably associated with manual restraint of the animal during vaccination. In addition, none of the animals had any evidence of virus shedding as the control animals remained sero-negative for antibodies to the vaccine virus while being confined in the same pens with vaccinated animals and this further confirmed the safety of the vaccine and the vaccination device. The safety results are consistent with those of previous studies conducted in Tanzania and USA involving the use of the RVFV arMP-12ΔNSm21/384 vaccine in sheep, calves and goats [57,62].
Neutralizing antibody is considered to be the main protective arm of the immune response against RVFV infection. Currently, the minimal standard PRNT80 protective titer for at risk personnel working with RVFV is ≥ 1:40 [26]. In this study, neutralizing antibody of all vaccinated goats, sheep and calves was first detected starting from either day 5 to 14 PV with titers ranging from 1:10 to 1:40 similar to what was observed in previously studies done in sheep and calves [57]. Seroconversion of two sheep (#930 and #935) vaccinated with 100 μl of the vaccine dose was detected on the same days as those vaccinated with 50 μl and antibody titers were higher in the two sheep vaccinated with 100 μl as compared to those vaccinated with 50 μl. The results are in agreement with those observed in previous sheep study [62], whereby animals were given 1 ml 1 × 105 PFU of the same vaccine dose and their antibody titers increased to as high as ≥ 1:160. Furthermore the results for 100 μl (8 × 105 PFU) vaccine dose in sheep were similar to those observed in a rhesus monkey study [53,58]. This demonstrated that the the IN vaccine dose affects the immune response of the animals regardless of the animal species, and that a higher dose may be needed than that used with needle delivery.
Most of the sheep, goats and calves that responded to the vaccine had antibody titers of ≥ 1:40 which has been the target end point of vaccine titers in large mammals. Lower titers have been found to be protective against virulent RVF virus in rodent studies. Goats developed the highest antibody response as compared to sheep and calves and the results were similarly to what was observed by others on testing the safety of the clone 13 vaccine [59]. Some of the goats, sheep and calves vaccinated with arMP-12ΔNSm21/384 vaccine did not have any detectable antibody titers by PRNT80, displaying individual variation among group of the animals. Therefore, regardless of the routes and doses, antibody response always can be affected by factors such as genetics and animal health status. Also, a variable in administering vaccines via the mucosal route is the inefficient uptake of the vaccine that can vary from animal to animal [60]. The data shown here support using the route for RVFV vaccines.
The study was planned to evaluate a vaccine dose volume of 50 μl. The results showed that antibody titers were higher for animals vaccinated with 100 μl of the vaccine dose than those vaccinated with 50 μl, though neutralization antibody titers produced from both dose amounts were mostly at the protective level. The 100 μl dose results were comparable to those observed from previous studies done in USA and Africa [57,59,61,62] and also similar to study done in Canada, that also revealed a titer of ≥ 1:40 is protective [63]. Possible differences in antibody responses elicited in animals vaccinated IN with 50 μl as compared to those vaccinated with 100 μl reflected differences in the amount of vaccine dose. From the results of this study, antibody titers of 50 μl injected animals are believed to be lower because of the low dosage used (4 × 105 PFU). The evaluation of the RVFV arMP-12ΔNSm21/384 vaccine using the IN route of vaccination was a proof of concept study and the first known study conducted in livestock using this route of vaccination.
RVFV vaccine delivery via the IN route offers a promising alternative to classical parenteral route since the route because it removes the complications of needle delivery. Also, the IN route has been shown to be more advantageous than parenteral route as it offers protective mucosal and systemic immunity unlike the invasive route that provides systemic immunity and weak mucosal immunity that may not provide more robust protection against RVF [64-69].
As a proof of concept study, the volume of the RVFV arMP- 12ΔNSm21/384 vaccine dose was 50 μl but contained almost the same concentration (4 × 105 PFU) as the maximum dose used in most other studies [57,62,63,69,70]. The lower volume was used because all the available intranasal doses packaged in the Mystic unit dose blisters were designed to deliver 50 μl of the volume. Thus, the 50 μl volume of the vaccine allowed for the determination if the lower volume would elicit an immune response In case the lower volume did not elicit an immune response, 2 sheep were vaccinated with 100 μl or twice the 50 μl volume administered to the other animals to gain a preliminary understanding of the volume required eliciting an immune response. The findings revealed that sheep vaccinated with 8 x 105 PFU of (double volume) of the RVFV arMP-12ΔNSm21/384 vaccine resulted in a higher immune response, thus demonstrating that the vaccine dose was a key determinant of the extent of the immune response [71].
There is much interest in the development of vaccines for mucosal application, and an attenuated immunogen given by a mucosal route that will elicit a robust and protective immune response against both parenteral and aerosol infection would be highly desirable [46,64].
Conclusion and Recommendations
These data support intranasal vaccination of livestock using RVFV arMP-12ΔNSm21/384 was safe and immunogenic. The response was dose dependent since the 100 μl dose of vaccine provided a higher antibody response than the 50 μl dosage. The needle-free non-invasive nasal route is advantageous for vaccination because it can be given by lay personnel with minimal training and therefore reduces the need to be administered by specialised workers. The administration of a vaccine by a noninvasive route would have advantages; especially in the field in remote areas where the ability to maintain sterility of hypodermic equipment is difficult or when large numbers of persons must be given the vaccine in a post event scenario. This is the first study that tested the safety and immunogenicity of the arMP-12ΔNSm21/384 vaccine via IN route in African ruminants. However, more studies must be done to determine the optimum dose of the IN route in a large number of domestic African ruminants. Also, field studies should be done to test the safety and immunogenicity of this vaccine, especially to determine the duration of the antibody titers and to determine reproductive safety of the arMP-12ΔNSm21/384 vaccine in pregnant ewes.
Acknowledgement
Many thanks to Dr. Mhando, Peter Marwa and Shida Mkuya for their tireless support of handling and maintaining animals throughout the study. Similarly, sincere thanks to the United States Agency for International Development (USAID) under Feed the Future Innovation Laboratory for Rift Valley Fever Control in Agriculture for providing the scholarship and funding for this study. Also, special appreciation goes to Sokine University of Agriculture virology laboratory team and University of Texas at El Paso (UTEP) team for their cooperation and assistance during the study.
Conflict of Interest
All authors declare no conflict of interest.
Author´s Contributions
The authors, DMW, GEB, PW, MM, and SB conceived and designed the experiments. The experiments were performed by SB, EA, LS, PP, JM, LM, TS and MM, and the same authors participated in the acquisition, analysis, and interpretation of the data for the work. The authors, SB, EA, PP, DMW, PW, GEB, MM, TS and JM prepared the manuscript.
Support
This study was funded under a subcontract from the University of Texas at El Paso (UTEP), Texas and the recipient of a Cooperative Agreement (AID-OAA-A-13-00084) from the USAID.
Disclaimer
The author’s views expressed in this publication do not necessarily reflect the views of the USAID or the United States Government. The contents are the responsibility of the Department of Microbiology, Parasitology and Biotechnology, Sokoine University of Agriculture, Morogoro, Tanzania and do not necessarily reflect the views of UTEP, USAID or the United States Government. The funders did not have any role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
REFERENCES
- Daubney R, Hudson J. R, Garnham P. C. Enzootic hepatitis or rift valley fever. An undescribed virus disease of sheep cattle and man from east africa. J Pathol Microbiol. 1931;34(4) :545–579.
- Kamal SA. Observations on rift valley fever virus and vaccines in Egypt. Virol J. 2011;8(1):1-9.
- Chevalier V, Thiongane Y, Lancelot R. Endemic transmission of rift valley fever in Senegal. Transbound Emerg Dis. 2009 ;56(9‐10):372-374.
- Sang R, Lutomiah J, Said M, Makio A, Koka H, Koskei E, et al. Effects of irrigation and rainfall on the population dynamics of rift valley fever and other arbovirus mosquito vectors in the epidemic-prone Tana River County, Kenya. J Med Entomol. 2017;54(2):460-470.
- Taylor D, Hagenlocher M, Jones AE, Kienberger S, Leedale J, Morse AP. Environmental change and Rift Valley fever in eastern Africa: Projecting beyond healthy futures. Geospat Health. 2016;11:115-128.
- Madani TA, Al-Mazrou YY, Al-Jeffri MH, Mishkhas AA, Al-Rabeah AM, Turkistani AM, et al. Rift valley fever epidemic in Saudi Arabia: Epidemiological, clinical, and laboratory characteristics. Clin Infect Dis. 2003;37(8):1084-1092.
- Redding DW, Tiedt S, Lo Iacono G, Bett B, Jones KE. Spatial, seasonal and climatic predictive models of Rift Valley fever disease across Africa. Philos Trans R Soc Lond B Biol Sci. 2017;372:1–9.
- Al-Afaleq AI, Elzein EM, Mousa SM, Abbas AM. A retrospective study of Rift valley fever in Saudi Arabia. PLoS Negl Trop Dis. 2003;22(3): 867-872.
- Caminade C, Ndione JA, Diallo M, MacLeod DA, Faye O, Ba Y, et al. Rift valley fever outbreaks in Mauritania and related environmental conditions. Int J Environ Res Public Health. 2014;11(1):903-918.
- Glancey MM, Anyamba A, Linthicum KJ. Epidemiologic and environmental risk factors of Rift valley fever in southern Africa from 2008 to 2011. Vector Borne Zoonotic Dis. 2015;15(8):502-511.
- Himeidan YE, Kweka EJ, Mahgoub MM, El Rayah EA, Ouma JO. Recent outbreaks of rift valley fever in east africa and the middle east. Front Public Health. 2014;2:169.
- Jost CC, Nzietchueng S, Kihu S, Bett B, Njogu G, Swai ES, et al. Epidemiological assessment of the Rift valley fever outbreak in Kenya and Tanzania in 2006 and 2007. Am J Trop Med Hyg. 2010; 8365-8372.
- OIE Terrestrial Manual. Aetiology Epidemiology Diagnosis Prevention and Control References. OIE Terrestrial Manual. 2012 :1–5.
- Dar O, McIntyre S, Hogarth S, Heymann D. Rift Valley fever and a new paradigm of research and development for zoonotic disease control. Emerg Infect Dis. 2013;19(2):189.
- Kahlon SS, Peters CJ, LeDuc J, Muchiri EM, Muiruri S, Njenga MK, et al. Severe rift valley fever may present with a characteristic clinical syndrome. Am J Trop Med Hyg. 2010; 82(3): 371-375.
- Williams R, Malherbe J, Weepener H, Majiwa P, Swanepoel R. Anomalous high rainfall and soil saturation as combined risk indicator of rift valley fever outbreaks, South Africa, 2008-2011. Emerg Infect Dis. 2016; 22(12): 2054.
- Pepin M, Bouloy M, Bird BH, Kemp A, Paweska J. Rift Valley fever virus (Bunyaviridae: Phlebovirus): an update on pathogenesis, molecular epidemiology, vectors, diagnostics and prevention. Vet Res. 2010; 41(6): 61.
- Ikegami T, Makino S. The pathogenesis of rift valley fever. Viruses. 2011;3(5):493-519.
- Miller MM, Bennett KE, Drolet BS, Lindsay R, Mecham JO, Reeves WK, et al. Evaluation of the efficacy, potential for vector transmission, and duration of immunity of MP-12, an attenuated Rift Valley fever virus vaccine candidate, in sheep. J Clin Invest. 2015; 22(8): 930-937.
- Centers for Disease Control. Rift Valley Fever (RVF) Fact Sheet. National Center for Emerging and Zoonotic Infectious Diseases. 2000 ; 1–2.
- Mohamed M, Mosha F, Mghamba J, Zaki SR, Shieh WJ, Paweska J, et al. Epidemiologic and clinical aspects of a rift valley fever outbreak in humans in Tanzania, 2007. Am J Trop Med Hyg. 2010 ; 83: 22-27.
- Ross TM, Bhardwaj N, Bissel SJ, Hartman AL, Smith DR. Animal models of Rift Valley fever virus infection. Virus Res. 2012; 163(2): 417-423.
- Taylor LH, Latham SM. Woolhouse MEJ. Risk factors for human disease emergence. Phil Trans R Soc. Lond B. 2001; 356: 983-989.
- Gowtage-Sequeria S, Woolhouse ME. Host range and emerging and reemerging pathogens. Emerging Infectious Diseases. 2005; 11(12): 1842-1847.
- Reed C, Lin K, Wilhelmsen C, Friedrich B, Nalca A, Keeney A, et al. Aerosol exposure to rift valley fever virus causes earlier and more severe neuropathology in the murine model, which has important implications for therapeutic development. PLoS Negl Trop Dis. 2013; 7(4): e2156.
- Pittman PR, McClain D, Quinn X, Coonan KM, Mangiafico J, Makuch RS, et al. Safety and immunogenicity of a mutagenized, live attenuated Rift Valley fever vaccine, MP-12, in a Phase 1 dose escalation and route comparison study in humans. Vaccine. 2016; 34(4): 424-429.
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2018 Release [Internet]. Washington:ICTV; 2018.
- Huiskonen JT, Överby AK, Weber F, Grünewald K. Electron cryo-microscopy and single-particle averaging of Rift Valley fever virus: Evidence for GN-GC glycoprotein heterodimers. J Virol. 2009; 83(8): 3762-3769.
- Gauliard N, Billecocq A, Flick R, Bouloy M. Rift Valley fever virus noncoding regions of L, M and S segments regulate RNA synthesis. Virology. 2006; 351(1): 170-179.
- Liu L, Celma CC, Roy P. Rift valley fever virus structural proteins: Expression, characterization and assembly of recombinant proteins. J Virol. 2008; 5(1): 82.
- Raymond DD, Piper ME, Gerrard SR, Smith JL. Structure of the rift valley fever virus nucleocapsid protein reveals another architecture for RNA encapsidation. Proceedings of the National Academy of Sciences. 2010; 107(26): 11769-11774.
- Lara E, Billecocq A, Leger P, Bouloy M. Characterization of wild-type and alternate transcription termination signals in the rift valley fever virus genome. J Virol. 2011; 85(23): 12134-12145.
- Bird BH, Albariño CG, Hartman AL, Erickson BR, Ksiazek TG, Nichol ST. Rift valley fever virus lacking the NSs and NSm genes is highly attenuated, confers protective immunity from virulent virus challenge, and allows for differential identification of infected and vaccinated animals. J Virol. 2008; 82(6): 2681-2691.
- Barzon L, Lavezzo E, Costanzi G, Franchin E, Toppo S, Palù G. Next-generation sequencing technologies in diagnostic virology. J Clin Virol. 2013; 58(2): 346-350.
- Faburay B, LaBeaud AD, McVey DS, Wilson WC, Richt JA. Current status of rift valley fever vaccine development. Vaccines. 2017; 5(3): 29.
- Hunter P, Erasmus BJ, Vorster JH. Teratogenicity of a mutagenised Rift Valley fever virus (MVP 12) in sheep.
- Makoschey B, van Kilsdonk E, Hubers WR, Vrijenhoek MP, Smit M, Wichgers Schreur PJ, et al. Rift valley fever vaccine virus clone 13 is able to cross the ovine placental barrier associated with foetal infections, malformations, and stillbirths. PLoS Negl Trop Dis. 2016; 10(3): e0004550.
- McElroy AK, Albariño CG, Nichol ST. Development of a RVFV ELISA that can distinguish infected from vaccinated animals. J Virol. 2009; 6(1): 125.
- Morrill JC, Jennings GB, Caplen H, Turell MJ, Johnson AJ. Pathogenicity and immunogenicity of a mutagen-attenuated rift valley fever virus immunogen in pregnant ewes. Army medical research inst of infectious diseases fort detrick md. 1987.
- Morrill JC, Carpenter L, Taylor D, Ramsburg HH, Quance J, Peters CJ. Further evaluation of a mutagen-attenuated Rift Valley fever vaccine in sheep. Vaccine. 1991; 9(1): 35-41.
- Bird BH, Ksiazek TG, Nichol ST, MacLachlan NJ. Rift valley fever virus. J Am Vet Med Assoc. 2009; 234(7): 883-893.
- Ikegami T, Won S, Peters CJ, Makino S. Rescue of infectious rift valley fever virus entirely from cDNA, analysis of virus lacking the NSs gene, and expression of a foreign gene. J Virol. 2006; 80(6): 2933-2940.
- Won S. Reverse genetics system and fucntion of NSM protein of Rift Valley fever virus (family Bunyaviridae, genus phlebovirus). 2020.
- Nyundo S, Adamson E, Rowland J, Palermo PM, Matiko M, Bettinger GE, et al. Safety and immunogenicity of rift valley fever MP-12 and arMP-12ΔNSm21/384 vaccine candidates in goats (Capra aegagrus hircus) from Tanzania. Onderstepoort J Vet Res. 2019; 86(1): 1-8.
- Morrill JC, Laughlin RC, Lokugamage N, Pugh R, Sbrana E, Weise WJ, et al. Safety and immunogenicity of recombinant Rift Valley fever MP-12 vaccine candidates in sheep. Vaccine. 2013; 31(3): 559-565.
- Holmgren J, Czerkinsky C. Mucosal immunity and vaccines. Nat Med. 2005;11(4):S45-S53.
- Kozlowski PA, Williams SB, Lynch RM, Flanigan TP, Patterson RR, Cu-Uvin S, et al. Differential induction of mucosal and systemic antibody responses in women after nasal, rectal, or vaginal immunization: Influence of the menstrual cycle. J Immunol. 2002; 169(1): 566-574.
- Neutra MR, Kozlowski PA. Mucosal vaccines: The promise and the challenge. Nat Rev Immunol. 2006; 6(2): 148-158.
- Giudice EL, Campbell JD. Needle-free vaccine delivery. Adv Drug Deliv Rev. 2006;58(1):68-89.
- Mitragotri S. Immunization without needles. Nat Rev Immunol. 2005; 5(12): 905-916.
- Read RC, Naylor SC, Potter CW, Bond J, Jabbal-Gill I, Fisher A, et al. Effective nasal influenza vaccine delivery using chitosan. Vaccine. 2005; 23(35): 4367-4374.
- Chen K, Cerutti A. Vaccination strategies to promote mucosal antibody responses. Immunity. 2010 Oct 29; 33(4): 479-91.
- Morrill JC, Peters CJ. Mucosal immunization of rhesus macaques with Rift Valley fever MP-12 vaccine. J. Infect. 2011; 204(4): 617-625.
- Chevalier V, Pépin M, Plee L, Lancelot R. Rift Valley fever-a threat for Europe? Euro Surveill. 2010;15(10):19506.
- Iranpour M, Turell MJ, Lindsay LR. Potential for canadian mosquitoes to transmit rift valley fever virus1. J Am Mosq Control Assoc. 2011; 27(4): 363-369.
- Turell MJ, Dohm DJ, Geden CJ, Hogsette JA, Linthicum KJ. Potential for stable flies and house flies (Diptera: Muscidae) to transmit rift valley fever virus1. J Am Mosq Control Assoc. 2010; 26(4): 445-448.
- Morrill JC, Laughlin RC, Lokugamage N, Wu J, Pugh R, Kanani P, et al. Immunogenicity of a recombinant Rift Valley fever MP-12-NSm deletion vaccine candidate in calves. Vaccine. 2013; 31(43): 4988-4994.
- Morrill JC, Peters CJ. Protection of MP-12–vaccinated rhesus macaques against parenteral and aerosol challenge with virulent Rift Valley fever virus. J Infect Dis. 2011; 204(2): 229-236.
- Njenga MK, Njagi L, Thumbi SM, Kahariri S, Githinji J, Omondi E, et al. Randomized controlled field trial to assess the immunogenicity and safety of rift valley fever clone 13 vaccine in livestock. PLoS Negl Trop Dis. 2015; 9(3): e0003550.
- Zaman M, Chandrudu S, Toth I. Strategies for intranasal delivery of vaccines. Drug Deliv Transl Res. 2013; 3(1): 100-109.
- Dungu B, Louw I, Lubisi A, Hunter P, von Teichman BF, Bouloy M. Evaluation of the efficacy and safety of the rift valley Fever Clone 13 vaccine in sheep. Vaccine. 2010; 28(29): 4581-4587.
- Nyundo S, Adamson E, Rowland J, Palermo PM, Matiko M, Bettinger GE, et al. Safety and immunogenicity of rift valley fever MP-12 and arMP-12ΔNSm21/384 vaccine candidates in goats (Capra aegagrus hircus) from Tanzania. Onderstepoort J Vet Res. 2019; 86(1): 1-8.
- Weingartl HM, Nfon CK, Zhang S, Marszal P, Wilson WC, Morrill JC, et al. Efficacy of a recombinant Rift Valley fever virus MP-12 with NSm deletion as a vaccine candidate in sheep. Vaccine. 2014; 32(20): 2345-2349.
- Van Ginkel FW, Nguyen HH, McGhee JR. Vaccines for mucosal immunity to combat emerging infectious diseases. Emerg Infect Dis. 2000; 6(2): 123.
- Kalveram B, Lihoradova O, Indran SV, Ikegami T. Using reverse genetics to manipulate the NSs gene of the rift valley fever virus MP-12 strain to improve vaccine safety and efficacy. J Vis Exp. 2011; 57: e3400.
- Kozlowski PA, Williams SB, Lynch RM, Flanigan TP, Patterson RR, Cu-Uvin S, et al. Differential induction of mucosal and systemic antibody responses in women after nasal, rectal, or vaginal immunization: influence of the menstrual cycle. J Exp Med. 2002; 169(1): 566-574.
- Linthicum KJ, Davies FG, Kairo A, Bailey CL. Rift valley fever virus (family Bunyaviridae, genus Phlebovirus). Isolations from Diptera collected during an inter-epizootic period in Kenya. Epidemiology & Infection. 1985; 95(1): 197-209.
- Mansfield KL, Banyard AC, McElhinney L, Johnson N, Horton DL, Hernández-Triana LM, et al. Rift valley fever virus: A review of diagnosis and vaccination, and implications for emergence in Europe. Vaccine. 2015; 33(42): 5520-5531.
- Morrill JC, Laughlin RC, Lokugamage N, Wu J, Pugh R, Kanani P, et al. Immunogenicity of a recombinant rift valley fever MP-12-NSm deletion vaccine candidate in calves. Vaccine. 2013; 31(43): 4988-4994.
- Nyundo S, Adamson E, Rowland J, Palermo PM, Matiko M, Bettinger GE, et al. Safety and immunogenicity of rift valley fever MP-12 and arMP-12ΔNSm21/384 vaccine candidates in goats (Capra aegagrus hircus) from Tanzania. Onderstepoort J Vet Res. 2019; 86(1): 1-8.
- Saks AM. Multiple predictors and criteria of job search success. J Vocat Behav. 2006; 68(3): 400-415.
Citation: Nyundo S, Adamson EK, Rowland J, Salekwa L, Palermo PM, Matiko MK, et al. (2020) Assessment of the Immunogenicity of a Novel Live Recombinant Rift Valley Fever arMP-12ΔNSm21/384 Vaccine Candidate Following Intranasal Vaccination of Goats, Sheep and Calves in Tanzania. J Vaccines Vaccin. S5:002.
Copyright: © 2020 Nyundo S, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.