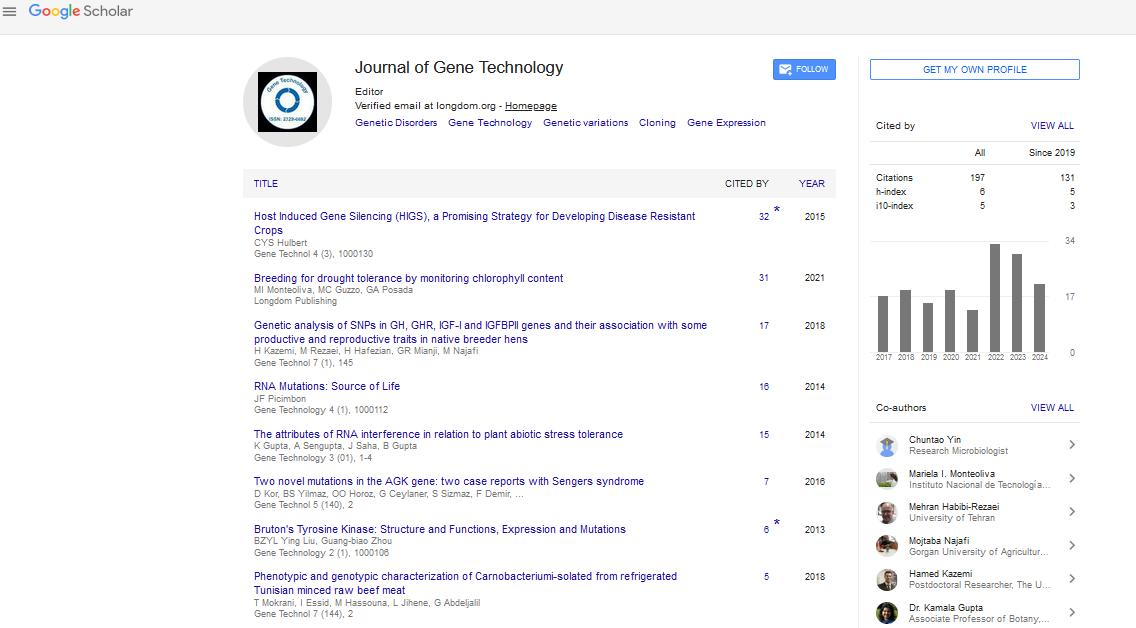

Indexed In
- Academic Keys
- ResearchBible
- CiteFactor
- Access to Global Online Research in Agriculture (AGORA)
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Review Article - (2020) Volume 9, Issue 2
A Possible Beneficial Effect of Sex Hormones on Vascular Aging through Expression of Sirtuin 1 Gene: A Narrative Review of the Literature
Takafumi Tsuchiya1*, Kenji Hara1, Kozo Takebayashi1, Koshi Hashimoto1 and Toshihiko Inukai22Department of Internal Medicine, Seibu General Hospital, Saitama 338-0824, Japan
Received: 17-Jun-2020 Published: 08-Jun-2020, DOI: 10.35248/2329-6682.20.9.150
Abstract
Sirtuin 1 (SIRT1) is a NAD+_dependent class III histone deacetylase, and a key gene linked to control of longevity, gene silencing, cell cycle progression, apoptosis, inflammation, stress resistance and energy homeostasis. SIRT1 is activated in response to low cellular energy stores and have been implicated in the control of many physiological processes including senescence. SIRT1 also regulates steroid hormone signaling through a variety of molecular mechanisms and modulate pathways that modify steroid hormone receptors. Declining of sex steroid hormones, including estrogens and androgens, is involved in the aging process and age-related diseases such as sarcopenia, falling, osteoporosis, cognitive and mood disorders, cardiovascular diseases, and sexual disturbance.
In this review, we will focus on the effects of sex steroid hormones on SIRT1 gene expression in endothelial cells and the advantages in the treatments with each hormone will be discussed in terms of understanding mammalian aging and longevity control.
Keywords
SIRT1; Estrogen; Androgen; Endothelial cells
Introduction
Sirtuin family
The sirtuins comprise a family of enzymes belonging to class III histone deacetylase (HDAC), which operate by removing acetyl groups from histones and other protein regulatory factors, resulting infunctional consequences on chromatin remodeling and gene expression profiles [1]. Moreover, the sirtuin family is highly conserved from archaebacteria to eukaryotes [2,3].
The life-prolonging effects of sirtuins were first described in yeast [4]. It was also shown that yeast sirtuin, silent information regulator 2 (Sir2), deficiency caused the short life span [5]. Sir2 is a NAD+_dependent histone deacetylase and its activity accounts for silencing, recombination suppression and extension of life span in vivo [4]. The mammalian sirtuin family consists of seven HDACs, sirtuin 1 (SIRT1) through sirtuin 7 (SIRT7), that share a conserved catalytic core domain and are expressed ubiquitously [6]. SIRT1, 6 and 7 localize to the nucleus, whereas SIRT3, 4 and 5 locate in mitochondria, and SIRT2 is primarily cytoplasmic [7]. Of seven mammalian proteins homologous to Sir2, SIRT1 and SIRT6 have been reported to be involved with longevity [8-10]. Sirtuins are activated in response to low cellular energy stores and have been implicated in the control of many physiological processes including senescence [11].
Sirtuin 1 (SURT1) and longevity
Energy metabolism is deeply involved in cellular aging. It is widely known that calorie restriction has the effect of suppressing aging and extending life span across species [12,13]. In particular, moderate calorie restriction in humans ameliorates multiple metabolic and hormonal factors that are implicated in the pathogenesis of type 2 diabetes, cardiovascular diseases and cancer, that are the leading causes of morbidity, disability and mortality [13]. Sirtuin 1 is a key gene linked to control of longevity, gene silencing, cell cycle progression, apoptosis, inflammation, stress resistance and energy homeostasis [14-19]. Sirtuin 1 transcription is activated during fasting, and triggers changes in metabolism that switches from gluconeogenesis to fat mobilization and fatty acid oxidation when fasting is prolonged [20]. Transgenic mice with whole-body overexpression of SIRT1 display phenotypes with metabolic features that protect against disorders associated with diet-induced obesity such as metabolic syndrome, type 2 diabetes and liver steatosis, although these effects are not sufficiently potent to affect longevity [21-23]. However, brain-specific SIRT1-overexpressing (BRASTO) transgenic mice show significant life span extension in both males and females, and aged BRASTO mice exhibit phenotypes consistent with a delay in aging [8]. These phenotypes are mediated by enhanced neural activity specifically in the dorsomedial and lateral hypothalamic nuclei (DMH and LH, respectively), indicating the importance of DMH/LHpredominant SIRT1 activity in theregulation of aging and longevity in mammals. Recently, adipose-tissue-specific overexpression of nicotinamide phosphoribosyl transferase (NAMPT) mice exhibit significant extension of median lifespan and delay in aging [24]. In mammals, NAMPT is the ratelimiting enzyme in a major NAD+ biosynthetic pathway, an essential substrate for SIRT1 activity [25]. It is suggested that the mammalian NAD+_dependent protein deacetylase SIRT1 and the key NAD+_biosynthetic enzyme NAMPT mediate an intertissue communications [26]. Although the relationship between SIRT1 and human life span is still controversial [27], there are a few reports that SIRT1 polymorphisms may be associated with body mass index and long-term weight changes [28,29].
Sex Hormones And Aging
SIRT1 also regulates steroid hormone signaling through a variety of molecular mechanisms and modulate pathways that modify steroid hormone receptors [30]. Declining of sex steroid hormones, including estrogens and androgens, is involved in the aging process and age-related diseases such as sarcopenia, falling, osteoporosis, cognitive and mood disorders, cardiovascular diseases, and sexual disturbance [31]. Estrogen- and testosteronedependent actions play a vital role in mitochondrial process implicated in aging [32]. The incidence of cardiovascular and vascular diseases is greater in men compared with age-matched premenopausal women [33]. Reduction of endogenous estrogen levels increases risk of bone fracture, cardiovascular diseases and Alzheimer's disease (AD) in postmenopausal women [34]. Therefore, during menopause the incidence in women dramatically increases, supporting a long-standing hypothesis that estrogens might provide vascular protection [35]. However, studies showed that estrogen therapy plays osteoprotective roles in both osteoporotic humans and rodents [36], while whether estrogen therapy can protect against heart diseases or AD remains controversial [37]. Estrogen reduces the risk of developing atherosclerosis in premenopausal women, whereas in post-menopausal women who may have established atherosclerotic diseases, estrogen increases the risk of myocardial diseases through the effects on plaque stability and clot formation [38]. Estrogens may also modulate cardiovascular health through expression of SIRT1, possibly in the AKT and ERK signal pathways [39]. It is also known that SIRT1 is the binding partner of estrogen receptor α ( ER α ) in mammary epithelial cells and that the ER-SIRT1 complex functions as a transcriptional activator of superoxide dismutase (SOD) and glutathione peroxidase (Gpx) and as a transcriptional repressor of p53 and cyclin G2, whereas ERα bound to the promoter for SIRT1 and increased its transcription in breast cancer cells [40].
On the other hand, it has been known that a decrease in androgens, particularly testosterone, as a result of aging in men or bilateral ovariectomy in women, is associated with hypertension, diabetes, and atherosclerosis and that testosterone replacement therapy may benefit these people [41], although there is evidence that androgen use has been associated with premature coronary diseases in athletes and impaired vascular reactivity in female-to-male trans-sexuals [42]. Studies have also shown alterations in mood, libido, and cognition resulting from testosterone deficiency [43]. Dehydroepiandrostendione (DHEA) may have beneficial effects that have been shown in vitro and in vivo, including the stimulation of immunity and then the suppression of diabetes, atherosclerosis, dementia, obesity and osteoporosis [44]. Delineating hormonal signaling changes that occur across a lifespan and searching interventions may improve the quality of life (QOL) of elderly people and extend longevity [45].
Sirt1 And Vascular Aging
Atherogenic stimuli, including diabetes, dyslipidemia, and oxidative stress, induce vascular dysfunction, leading to atherosclerosis, which is a key pathological basis for cardiovascular diseases such as ischemic heart diseases and strokes [46]. SIRT1 is highly expressed in human vascular endothelial cells [47] and has a potential antioxidative stress activity in vascular endothelial cells. The inhibition of SIRT1 with pharmacological agents or siRNA leads to an elevation of reactive oxygen species (ROS) levels in an animal model [48]. In human umbilical vein endothelial cells (HUVEC), SIRT1 inhibition increased p53 acetylation and induced premature senescence-like phenotype in parallel with increased plasminogen activator inhibitor-1 (PAI-1) and decreased endothelial nitric oxide synthase (eNOS) expression, whereas overexpression of SIRT1 deranged expression of PAI-1 and eNOS and reversed premature senescence induced by oxidative stress [49]. Those data have indicated a definite relationship between SIRT1 and ROS. The signaling networks of SIRT1 involved in ROS resistance include SIRT1/Forkhead Box class O transcription factors (FOXOs), SIRT1/Nuclear Factor - kB (NF -kB), SIRT1/NADPH oxidase (NOX), SIRT1/SOD, and SIRT1/eNOS pathways [50]. A previous study demonstrated that calorie restriction increased SIRT1 in the white adipose tissue of wild type mice, and this effect was abolished in eNOSknockout mice [51], suggesting that ROS might control SIRT1 expression as well.
Vascular Aging And Sex Hormones
It has been reported that ERα expression modulated by estrogen in endothelial cells is related to eNOS activation [52]. Another report described that ERβ expression in the endothelium was reduced in aging mice, and the expression of ERα and SIRT1 was not changed, while SIRT1 activity was declined [53]. Estrogens include estrone (E1), estradiol (E2), and estriol (E3). E2 partially suppressed angiotensin II-induced contractions, restored the protein expression of SIRT1/P-AMPK and suppressed histone H3 acetylation in aorta of a post- menopausal metabolic syndrome model induced in ovariectomized rats by feeding a high-fat diet [54]. Recently, it is reported that either E2 or selective estrogen receptor modulator (SERM) administration increased SIRT1 in endothelial cells and activated eNOS, resulting in decreased vascular senescence and atherosclerotic lesions [55]. The effect of SERM on upregulating SIRT1 was abolished in eNOS-knockout underwent ovariectomy (OVX) mice, OVX+SERM mice treated with a NOS inhibitor also showed no differences in arterial SIRT1 expression and senescence, suggesting that SIRT1 expression is regulated by estrogen-induced eNOS activation. Treatment of human endothelial cells with E2 has been reported to induce SIRT1 gene expression [55,56]. In our own study, treatmentof human aortic endothelial cells (HAEC) with E1 showed similar effects of E2 treatment, while E3 failed to induce SIRT1 gene expression [56]. The beneficial effects of E2 appear to occur from some properties including antioxidant and to imply an overall anti-aging action [57]. On the other hand, transvaginal E3 potentially offers a suitable physiologic delivery as a clinical benefit, however, E3 might be a weak activator of longevity, in contrast to E1 and E2 [58]. Then, the E1- and E2-induced SIRT1 expression was not diminished by high glucose levels [56]. It is consistent with a previous finding that control of oxidative stress by AMPK activation or antioxidants could restore normal estrogen responses, even in hyperglycemia [59], suggesting a possible benefit of anti-atherogenic effects of estrogens in female patients with diabetes.
Androgen receptor (AR) is also expressed in endothelial cells [35]. Epidemiological and clinical data have indicated that androgens are independent factors that contribute to the higher male susceptibility to atherosclerosis through adverse effects on lipids, blood pressure, and glucose metabolism [41,60].
A retrospective national cohort study showed the association of testosterone therapy with mortality, myocardial infarction, and stroke in men with low testosterone levels [61], indicating that excessive testosterone exposure may be detrimental to the cardiovascular system [62-64]. There are a few reports to investigate the effect of androgens on SIRT1 expression in endothelial cells. In human umbilical vein endothelial cells (HUVEC), it is shown that oxidative stress decreases eNOS and SIRT1 and increases PAI-1 expression, and dihydrotestosterone (DHT) or testosterone treatment prevented these changes and increases the phosphorylation of eNOS at Ser1177 [65]. Another report evaluated the endothelial function of the corpus cavernosum in Otsuka Long-Evans Tokushima Fatty (OLETF) rats, which recapitulate type 2 diabetes [66]. The expression of eNOS and Sirt1 mRNA was decreased and that of inducible NO synthase (iNOS), IL-6, and TNF-mRNA was increased, while androgen replacement therapy improved the mRNA expression in OLETF rats. They concluded that androgen replacement therapy suppressed inflammation in rats with type 2 diabtetes and metabolic disorders and improved their endothelial and erectile functions. In our own study, treatments of HAEC with testosterone and DHEA have been also shown to induce SIRT1 mRNA expression, and those effects were not inhibited under culture condition with high glucose levels [56]. However, treatment with androstenedione exerted little effects on SIRT1 mRNA expression in HAEC, which is consistent with a report showing that androstenedione had no effects on development of bones, including sternebrae and soft tissues [67]. Androstenedione is a metabolite of the androgen pathway, suggesting that it may have a relatively weak bioactivity in cultured cells (Table 1).
| Estrogens | Androgens | ||||||
|---|---|---|---|---|---|---|---|
| Condition | E1 | E2 | E3 | Testosterone | Androstenedione | DHT | DHEA |
| Normal glucose | ↑↑↑ | ↑↑↑ | → | ↑↑ | → | ↑↑ | ↑ |
| High glucose | ↑↑↑ | ↑↑↑ | → | ↑↑ | → | N. A. | ↑ |
N. A. indicates that data were not available; E1: Estrone; E2: Estradiol; E3: Estriol; DHT: Dihydrotestosterone; DHEA:Dehydroepiandrostendione.
Table 1: The effects of sex steroid hormones on SIRT1 expression in endothelial cells and tissues [54-56, 65,66].
Conclusion
We have reviewed the recent progresses related to SIRT1- mediated beneficial effects of sex steroid hormones on vascular aging and discussed the possibility of the sex hormone treatment on endothelial cells. However, sex hormones appear to be at a significantly increased risk to have or to develop specific cancers, although we did not mention it in this review. Thus, we should pay attention to both the advantages and disadvantages of sex hormone treatment and further understanding of the molecular mechanism of the protective effects of sex hormones against aging and to extend longevity should be required.
REFERENCES
- Milazzo G, Mercatelli D, Di Muzio G, Triboli L, De Rosa P, Perini G, et al. Histone deacetylases (HDACs): Evolution, specificity, role in transcriptional complexes, and pharmacological actionability. Genes (Basel). 2020;11(5):556.
- Frye RA. Characterization of five human cDNAs with homology to the yeast SIR2 gene: Sir2-like proteins (sirtuins) metabolize NAD and may have protein ADP-ribosyltransferase activity. Biochem Biophys Res Commun. 1999;260(1):273-279.
- Frye RA. Phylogenetic classification of prokaryotic and eukaryotic Sir2-like proteins. Biochem Biophys Res Commun. 2000;273(2):793-798.
- Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795-800.
- Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999;13(19):2570-2580.
- Michan S, Sinclair D. Sirtuins in mammals: Insights into their biological function. Biochem J. 2007;404(1):1-13.
- Michishita E, Park JY, Burneskis JM, Barrett JC, Horikawa I. Evolutionarily conserved and nonconserved cellular localizations and functions of human SIRT proteins. Mol Biol Cell. 2005;16(10):4623-4635.
- Satoh A, Brace CS, Rensing N, Cliften P, Wozniak DF, Herzog ED, et al. Sirt1 extends life span and delays aging in mice through the regulation of Nk2 homeobox 1 in the DMH and LH. Cell Metab. 2013;18(3):416-430.
- Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, Gellon L, et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006;124(2):315-29.
- Kanfi Y, Naiman S, Amir G, Peshti V, Zinman G, Nahum L, et al. The sirtuin SIRT6 regulates lifespan in male mice. Nature. 2012;483(7388):218-221.
- Winnik S, Auwerx J, Sinclair DA, Matter CM. Protective effects of sirtuins in cardiovascular diseases: from bench to bedside. Eur Heart J. 2015; 36(48): 3404-3412.
- López-Lluch G, Navas P. Calorie restriction as an intervention in ageing. J Physiol. 2016;594(8):2043-2060.
- Most J, Tosti V, Redman LM, Fontana L. Calorie restriction in humans: An update. Ageing Res Rev. 2017;39:36-45.
- Dali-Youcef N, Lagouge M, Froelich S, Koehl C, Schoonjans K, Auwerx J. Sirtuins: the 'magnificent seven', function, metabolism and longevity. Ann Med. 2007;39(5):335-345.
- Greiss S, Gartner A. Sirtuin/Sir2 phylogeny, evolutionary considerations and structural conservation. Mol Cells. 2009;28(5):407-415.
- Haigis MC, Guarente LP. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006;20(21):2913-2921.
- Yamamoto H, Schoonjans K, Auwerx J. Sirtuin functions in health and disease. Mol Endocrinol. 2007;21(8):1745-1755.
- Yang T, Fu M, Pestell R, Sauve AA. SIRT1 and endocrine signaling. Trends Endocrinol Metab. 2006;17(5):186-191.
- Brunet A, Sweeney LB, Sturgill JF, Chua KF, Greer PL, Lin Y, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303(5666):2011-2015.
- Pardo PS, Boriek AM. SIRT1 Regulation in Ageing and Obesity. Mech Ageing Dev. 2020;188:111249.
- Bordone L, Cohen D, Robinson A, Motta MC, van Veen E, Czopik A, et al. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell. 2007;6(6):759-767.
- Pfluger PT, Herranz D, Velasco-Miguel S, Serrano M, Tschöp MH. Sirt1 protects against high-fat diet-induced metabolic damage. Proc Natl Acad Sci. 2008;105(28):9793-9798.
- Herranz D, Muñoz-Martin M, Cañamero M, Mulero F, Martinez-Pastor B, Fernandez-Capetillo O, et al. Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat Commun. 2010;1:3.
- Yoshida M, Satoh A, Lin JB, Mills KF, Sasaki Y, Rensing N, et al. Extracellular Vesicle-Contained eNAMPT Delays Aging and Extends Lifespan in Mice. Cell Metab. 2019;30(2);329-342:e5.
- Imai S. Nicotinamide phosphoribosyltransferase (Nampt): a link between NAD biology, metabolism, and diseases. Curr Pharm Des. 2009;15(1):20-28.
- Imai SI. The NAD World 2. 0: the importance of the inter-tissue communication mediated by NAMPT/NAD. NPJ Syst Biol Appl. 2016;2:16018.
- Di Bona D, Accardi G, Virruso C, Candore G, Caruso C. Association between genetic variations in the insulin/insulin-like growth factor (Igf-1) signaling pathway and longevity: a systematic review and meta-analysis. Curr Vasc Pharmacol. 2014;12(5):674-681.
- Higashibata T, Wakai K, Naito M, Morita E, Hishida A, Hamajima N, et al. Effects of self-reported calorie restriction on correlations between SIRT1 polymorphisms and body mass index and long-term weight change. Gene. 2016;594(1):16-22.
- Zillikens MC, van Meurs JB, Rivadeneira F, Amin N, Hofman A, Oostra BA, et al. SIRT1 genetic variation is related to BMI and risk of obesity. Diabetes. 2009;58(12):2828-2834.
- Moore RL, Dai Y, Faller DV. Sirtuin 1 (SIRT1) and steroid hormone receptor activity in cancer. J Endocrinol. 2012;213(1):37-48.
- Samaras N, Papadopoulou MA, Samaras D, Ongaro F. Off-label use of hormones as an antiaging strategy: a review. Clin Interv Aging. 2014;9:1175-1186.
- Vasconsuelo A, Milanesi L, Boland R. Actions of 17β-estradiol and testosterone in the mitochondria and their implications in aging. Ageing Res Rev. 2013;12(4):907-917.
- Kannel WB. The Framingham study. Br Med J. 1976;2(6046):1255.
- Cui J, Shen Y, Li R. Estrogen synthesis and signaling pathways during aging: from periphery to brain. Trends Mol Med. 2013;19(3):197-209.
- Torres-Estay V, Carreño DV, San Francisco IF, Sotomayor P, Godoy AS, Smith GJ. Androgen receptor in human endothelial cells. J Endocrinol. 2015;224(3):R131-137.
- Frenkel B, Hong A, Baniwal SK, Coetzee GA, Ohlsson C, Khalid O, et al. Regulation of adult bone turnover by sex steroids. J Cell Physiol. 2010;224(2):305-310.
- Hulley S, Grady D, Bush T, Furberg C, Herrington D, Riggs B, et al. Randomized trial of estrogen plus progestin for secondary prevention of coronary heart disease in postmenopausal women. Heart and Estrogen/progestin Replacement Study (HERS) Research Group. JAMA. 1998;280(7):605-613.
- Hormones and cardiovascular health in women. Hum Reprod Update. 2006;12(5):483-97.
- Lee CH, Su SC, Chiang CF, Chien CY, Hsu CC, Yu TY, et al. Estrogen modulates vascular smooth muscle cell function through downregulation of SIRT1. Oncotarget. 2017;8(66):110039-110051.
- Elangovan S, Ramachandran S, Venkatesan N, Ananth S, Gnana-Prakasam JP, Martin PM, et al. SIRT1 is essential for oncogenic signaling by estrogen/estrogen receptor α in breast cancer. Cancer Res. 2011;71(21):6654-6664.
- Nheu L, Nazareth L, Xu GY, Xiao FY, Luo RZ, Komesaroff P et al. Physiological effects of androgens on human vascular endothelial and smooth muscle cells in culture. Steroids. 2011;76(14):1590-1596.
- Death AK, McGrath KC, Sader MA, Nakhla S, Jessup W, Handelsman DJ, et al. Dihydrotestosterone promotes vascular cell adhesion molecule-1 expression in male human endothelial cells via a nuclear factor-kappaB-dependent pathway. Endocrinology. 2004;145(4):1889-1897.
- Kaufman JM, Vermeulen A. The decline of androgen levels in elderly men and its clinical and therapeutic implications. Endocr Rev. 2005;26(6):833-876.
- Nawata H, Yanase T, Goto K, Okabe T, Ashida K. Mechanism of action of anti-aging DHEA-S and the replacement of DHEA-S. Mech Ageing Dev. 2002;123(8):1101-6.
- Diamanti-Kandarakis E, Papalou O, Kandaraki EA, Kassi G. Mechanisms In Endocrinology: Nutrition as a mediator of oxidative stress in metabolic and reproductive disorders in women. Eur J Endocrinol. 2017;176(2):R79-R99.
- Hirase T, Node K. Endothelial dysfunction as a cellular mechanism for vascular failure. Am J Physiol Heart Circ Physiol. 2012;302(3):H499-505.
- Potente M, Ghaeni L, Baldessari D, Mostoslavsky R, Rossig L, Dequiedt F, et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007;21(20):2644-2658.
- Hori YS, Kuno A, Hosoda R, Tanno M, Miura T, Shimamoto K, et al. Resveratrol ameliorates muscular pathology in the dystrophic mdx mouse, a model for Duchenne muscular dystrophy. J Pharmacol Exp Ther. 2011;338(3):784-794.
- Ota H, Akishita M, Eto M, Iijima K, Kaneki M, Ouchi Y. Sirt1 modulates premature senescence-like phenotype in human endothelial cells. J Mol Cell Cardiol. 2007;43(5):571-579.
- Zhang W, Huang Q, Zeng Z, Wu J, Zhang Y, Chen Z. Sirt1 Inhibits Oxidative Stress in Vascular Endothelial Cells. Oxid Med Cell Longev. 2017;2017:7543973.
- Nisoli E, Tonello C, Cardile A, Cozzi V, Bracale R, Tedesco L et al. Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science. 2005;310(5746):314-7.
- Gavin KM, Seals DR, Silver AE, Moreau KL. Vascular endothelial estrogen receptor alpha is modulated by estrogen status and related to endothelial function and endothelial nitric oxide synthase in healthy women. J Clin Endocrinol Metab. 2009;94(9):3513-3520.
- Kong D, Zhan Y, Liu Z, Ding T, Li M, Yu H, et al. SIRT1-mediated ERβ suppression in the endothelium contributes to vascular aging. Aging Cell. 2016;15(6):1092-1102.
- Bendale DS, Karpe PA, Chhabra R, Shete SP, Shah H, Tikoo K. 17-β Oestradiol prevents cardiovascular dysfunction in post-menopausal metabolic syndrome by affecting SIRT1/AMPK/H3 acetylation. Br J Pharmacol. 2013;170(4):779-795.
- Sasaki Y, Ikeda Y, Miyauchi T, Uchikado Y, Akasaki Y, Ohishi M. Estrogen-SIRT1 Axis Plays a Pivotal Role in Protecting Arteries Against Menopause-Induced Senescence and Atherosclerosis. J Atheroscler Thromb. 2020;27(1):47-59.
- Tsuchiya T, Takei A, Tsujikado K, Inukai T. Effects of androgens and estrogens on sirtuin 1 gene expression in human aortic endothelial cells. Saudi Med J. 2020;41(4):361-368.
- Kumar P, Taha A, Kale RK, Cowsik SM, Baquer NZ. Physiological and biochemical effects of 17β estradiol in aging female rat brain. Exp Gerontol. 2011;46(7):597-605.
- Ali ES, Mangold C, Peiris AN. Estriol: emerging clinical benefits. Menopause. 2017;24(9):1081-1085.
- Chakrabarti S, Davidge ST. High glucose-induced oxidative stress alters estrogen effects on ERalpha and ERbeta in human endothelial cells: Reversal by AMPK activator. J Steroid Biochem Mol Biol. 2009;117(4-5):99-106.
- Liu PY, Death AK, Handelsman DJ. Androgens and cardiovascular disease. Endocr Rev. 2003;24(3):313-340.
- Vigen R, O'Donnell CI, Barón AE, Grunwald GK, Maddox TM, Bradley SM, et al. Association of testosterone therapy with mortality, myocardial infarction, and stroke in men with low testosterone levels. JAMA. 2013;310(17):1829-1836.
- Blesson CS, Chinnathambi V, Hankins GD, Yallampalli C, Sathishkumar K. Prenatal testosterone exposure induces hypertension in adult females via androgen receptor-dependent protein kinase Cδ-mediated mechanism. Hypertension. 2015;65(3):683-690.
- Finkle WD, Greenland S, Ridgeway GK, Adams JL, Frasco MA, Cook MB, et al. Increased risk of non-fatal myocardial infarction following testosterone therapy prescription in men. PLoS One. 2014;9(1):e85805.
- Walsh JP, Kitchens AC. Testosterone therapy and cardiovascular risk. Trends Cardiovasc Med. 2015;25(3):250-257.
- Ota H, Akishita M, Akiyoshi T, Kahyo T, Setou M, Ogawa S, et al. Testosterone deficiency accelerates neuronal and vascular aging of SAMP8 mice: protective role of eNOS and SIRT1. PLoS One. 2012;7(1):e29598.
- Kataoka T, Hotta Y, Maeda Y, Kimura K. Assessment of androgen replacement therapy for erectile function in rats with type 2 diabetes mellitus by examining nitric oxide-related and inflammatory factors. J Sex Med. 2014;11(4):920-929.
- Sprando RL, Collins TF, Black TN, Olejnik N, Grundel E, Ruggles DI. Effects of androstenedione on in utero development in rats. Food Chem Toxicol. 2004;42(6):917-924.
Citation: Tsuchiya T, Inukai T, Hara K, Takebayashi K and Hashimoto K (2020) A Possible Beneficial Effect of Sex Hormones on Vascular Aging through Expression of Sirtuin 1 Gene: A Narrative Review of the Literature Gene Technol. 9:2. DOI: 10. 35248/2329-6682.20. 9.150.
Copyright: © 2020 Tsuchiya T, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.